Article Text

Statistics from Altmetric.com
Compromised growth in utero is associated with prematurity and complications after birth. It has been proposed that poor intrauterine growth not only contributes to increased morbidity and mortality during infancy but also has the potential to compromise adult health and wellbeing.1 Over the past decade, epidemiological studies in several countries have shown that size at birth and/or placental weight predict adult health and disease.1-3 It has also been proposed that maternal undernutrition at critical stages of gestation can affect fetal growth and body shape. These effects may be mediated in part by changes in placental growth, as some studies have suggested that fetal:placental weight ratio at birth is a predictor of adult disease.4 Fetal:placental weight ratio in healthy human pregnancies at term is approximately 5:1 but disease states can lead to alterations in this ratio. Intrauterine growth restriction is often accompanied by a light placenta in both humans and sheep,5 whereas the hydropic infant may have a large oedematous placenta6 and diabetic women produce a large infant and a heavy placenta.7
As recently stated by Leon,8 there is a large weight of evidence linking altered size at birth to adult cardiovascular disease. However, scepticism still persists as to the influence of maternal and/or fetal nutrition as a potential cause of adverse fetal growth.9 A major reason for this scepticism is that the findings from Barker's and related studies are largely based on epidemiological evidence. Attempts to confirm or refute Barker's hypothesis using data from human pregnancies have only made limited progress because long term experimental, and/or intervention studies, are difficult to undertake or ethically not possible. Consequently, investigators have used alternative human studies including prospective cohort studies or “experiments of nature” such as twinning. It is the aim of the present discussion to consider the potential problems of such approaches in view of related findings from complementary animal models.
Problems associated with prospective cohort studies
Human population based studies using geographically adjacent and economically comparable areas in the same decade would be predicted to produce complementary findings. This is not, however, always the case. Initial findings by Godfrey et al who examined maternal nutrition in 538 women in Southampton showed a negative relation between maternal energy intake and both placental and birth weights.10 This contrasts with the more recent publication by Mathews et al who studied 693 women in Portsmouth and found no significant effect of maternal nutrition on size at birth.11 Comparison of these two papers indicates a number of striking differences. In the study by Godfrey et al the cut off points for energy intake, for example, are ⩽2080, 2560, ⩾2560 kcal/day (⩽8.69, 10.7, ⩾10.7 MJ/day) compared with ⩽1855, 2204, ⩾2205 kcal/day (⩽7.75, 9.21, ⩾9.22 MJ/day) by Mathews et al. It is unclear how these arbitrary points were selected. An alternative analysis would be to use all the raw data points to determine potential relations between maternal nutrition and birth weight. If the methods used to assess maternal nutrient intake are robust and reproducible, it should be possible to combine data from both studies which would give a more powerful sample size. Comparison of the mean data sets published indicates there is overlap in maternal energy intake between studies but that there is an inconsistent impact of increasing energy on birth weight (fig 1). Interestingly, there is little difference in placental weight between studies.10 ,11
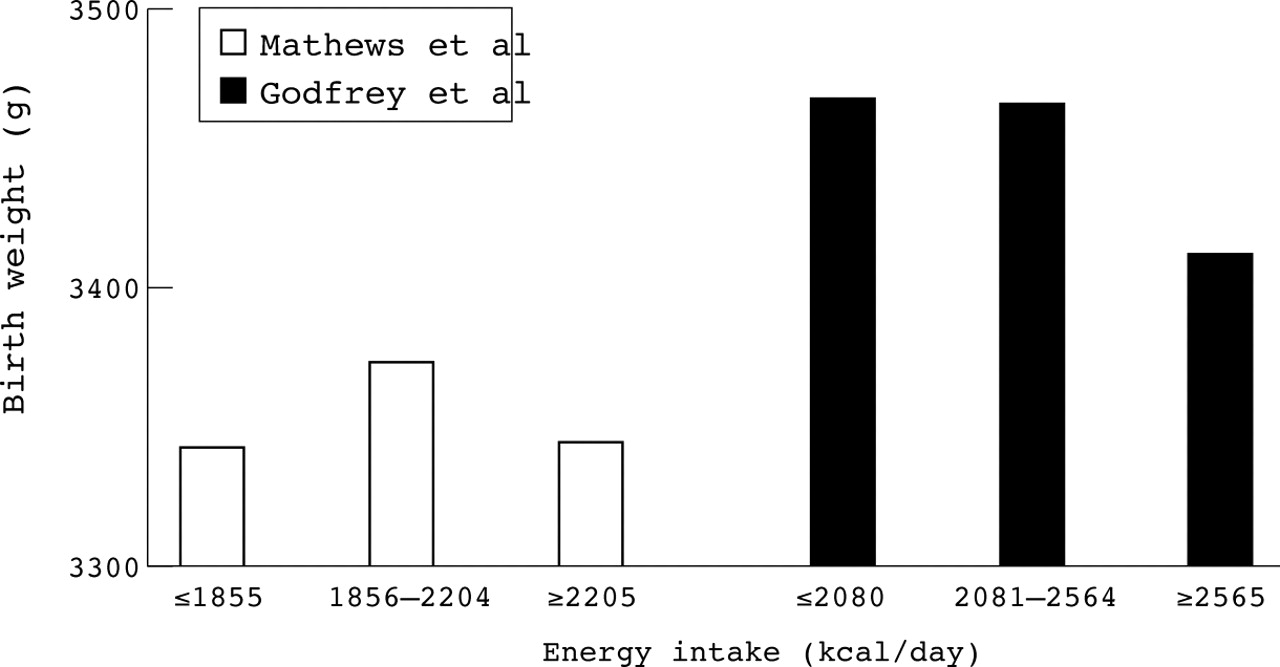
Animal studies showing the effect of maternal nutrition at specific stages of pregnancy on placental or fetal growth
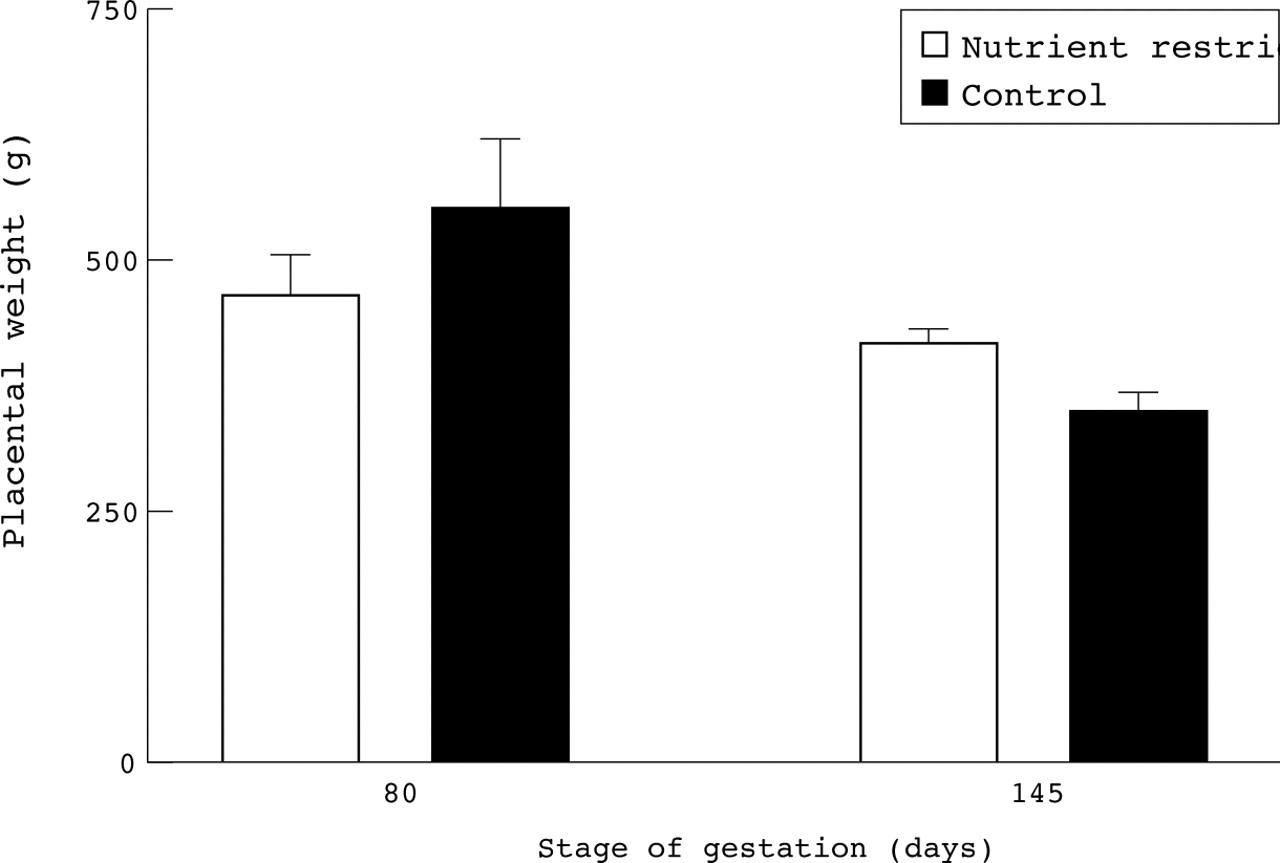
In the studies by both Mathews et aland Godfrey et al, maternal nutrition was estimated at only two points in gestation, with no account taken of changes in the intervening period, nor of maternal dietary intake with respect to dietary requirements. Studies in sheep have shown that such point sampling may miss important biological effects. Although undernutrition in early to mid gestation may initially restrict placental growth,12 if these ewes are subsequently adequately fed to fully meet the metabolisable energy requirements for late gestation, then a larger placenta results without any effect on fetal weight13 (fig 2). These findings are consistent with those of a recent analysis of the effects of maternal undernutrition on fetal and placental growth of infants born in 1944 to 1946 during the Dutch famine.14 This study showed that an energy intake of only 1300 kcal/day (5.43 MJ/day; about 56% of normal intake10), during the first trimester was associated with an increase in placental but not infant birth weight. In contrast, maternal undernutrition during the Dutch famine in late gestation was associated with a reduction in both placental and fetal weights.14 The fact that the reductions in food intake in the above animal study15 and period of famine14 were much greater than would be seen in contemporary UK pregnancies only serves to underline that the studies by Matthews et al and Godfreyet al may not be sensitive enough to show consistent effects of maternal nutrition on birth weight. Indeed, recent studies in sheep indicate that increasing maternal feed intake from adequate to excess of total energy requirements, enhances birth weight in the absence of any effect on placental weight (fig3).
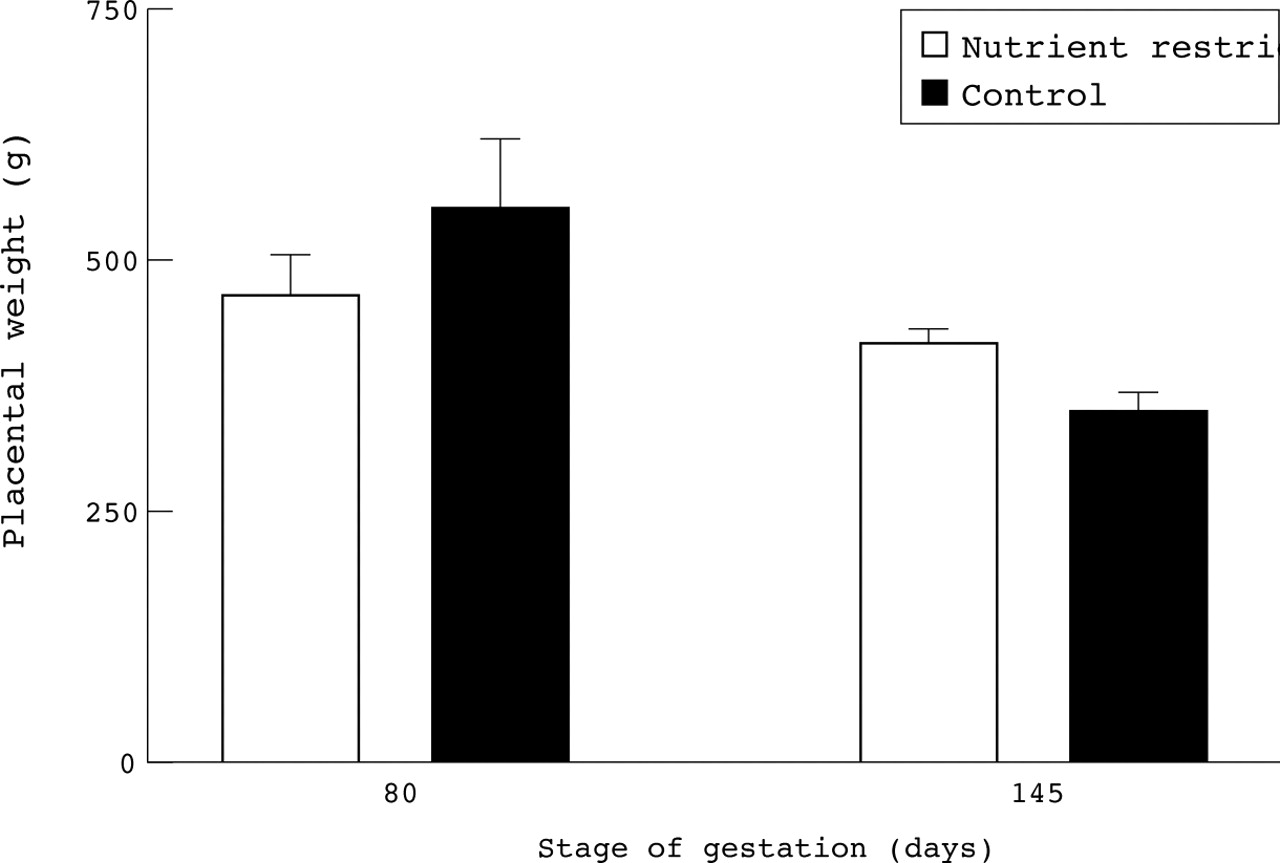
Effect of maternal nutrient restriction in early to mid gestation on placental weight at mid gestation (80 days gestation) or close to term (145 days gestation, with term being 147–8 days) in singleton bearing sheep. Nutrient restricted ewes consumed 3.71 MJ/day of metabolisable energy between 28 and 80 days gestation and 7.32 MJ/day between 80 and145 days gestation; controls consumed 6.49 MJ/day between 28 and 80 days and 7.11 MJ/day between 80 and 145 days. Values are means with their standard errors and were adapted from Clarke et al12 and Heasman et al.15

Effect of maternal nutritional enhancement between mid to late gestation (80 to 140–5 days gestation) on placental and fetal weights near to term (140–5 days gestation, with term being 147–8 days) in singleton bearing sheep. (A) Fetal weight. (B) Placental weight. Adequately fed ewes consumed 7.11 MJ/day and well fed ewes consumed 10.62 MJ/day, with all ewes being well fed from 28 to 80 days gestation. Values are means with their standard errors and were adapted from Budge et al,36 Dandrea et al,37 and Heasman et al.13
One consistent proposal by Barker has been that two specific groups of infants have increased vulnerability to hypertension and diabetes during adulthood, namely those that are either long and thin or short and fat at birth.1 ,4 ,16 Measurements in sheep have shown that the dimensions and endocrine status of lambs born to ewes that were nutrient restricted in early to mid gestation differ from controls. At term, fetuses of nutrient restricted ewes have a longer crown–rump length, height, and greater thoracic circumference.13
Poor fetal growth has been associated not only with an increased risk of adult cardiovascular disease but also insulin resistance.1 The permanent biological alterations which are thought to predispose to adult disease are termed fetal programming, although the mechanisms which link fetal growth retardation with adult pathologies have not been defined. The importance of insulin like growth factor I (IGF-I) in regulating fetal growth has been established from a number of direct and indirect studies. There are several reports in infants, calves, and lambs in which positive correlations between fetal body weight and plasma IGF-I concentration have been found.17-19 In fetal sheep, chronic (from 120 to 130 days gestation; term = 147 days) IGF-I infusion has been shown to promote skeletal maturation and growth of fetal liver, lungs, heart, kidneys, spleen, pituitary, and adrenal glands,20 as well as having anabolic effects on fetoplacental protein and carbohydrate metabolism.21 It is also established that chronic severe maternal undernutrition (25% of required energy and protein intake) in late gestation decreases both maternal and fetal plasma IGF-I concentrations in conjunction with a fall in fetal body and liver weights.22 In studies in which ewes were nutrient restricted between early to mid gestation, in contrast to controls, body dimensions of fetuses in nutrient restricted sheep do not correlate significantly with plasma IGF-I concentration in cord blood at term,15 indicative of loss of the normal relation between IGF-I and bone growth. Importantly, although there was a loss of correlation between fetal body dimensions and fetal plasma IGF-I concentration in fetuses from nutrient restricted fetuses, both plasma IGF-I concentrations and lamb birth weights were in the same range as controls.15 It is likely that even under experimental conditions in which birth weight is unaffected, the fetal somatotrophic axis is reset.
The extent to which maternal macro- or micronutrition may determine placental and fetal weights as well as size at birth remains to be fully determined and it is only by the combination of appropriate animal models in conjunction with more extensive population based studies that progress will be made. Each approach has strengths and weaknesses. Well defined experimental studies with the necessary controls can examine precise hypotheses but are usually limited by the small numbers involved and are often ethically impossible in humans. This contrasts with very large observational studies which have no controls, containing a range of confounders and relying on both food diaries and food frequency questionnaires that are not always validated in the same way.11 In the latter studies, absence of evidence does not always equate with evidence that a specific effect is absent. There is an increased chance of a type II statistical error if, for example, food diaries are imprecisely completed by individuals who are conscious that they are under- or overeating,23 with differences regressing towards the mean.
Problems associated with the “twin model” of fetal development
A further point to consider is the impact of fetal number. Recent studies24-26 have attempted to test Barker's hypothesis that undernutrition, manifest in slow growth and consequent low birth weight, programmes the fetus for higher blood pressure in later life. In one study, Williams and Poulton24 studied a cohort of children and measured systolic blood pressure at ages 9 and 18 years, relating this to potential constraints on fetal nutrition such as twinning, maternal smoking, and maternal size. It was hypothesised that systolic blood pressure would be higher in twin subjects than normal (singleton) controls. In contrast to the hypothesis, it was found that twinning was associated with a decrease in the average systolic blood pressure. However, their cohort of twins was only 2.8% of the total cohort and no comment was made on placental size or type (mono- or dichorionic).24 This is important as the authors assumed that fetal and placental growth and outcome in human twin pregnancies is similar to singleton pregnancies irrespective of whether the fetuses share a placenta (monochorionic) or had separate placentae (dichorionic), which is not the case.27 ,28 The regression coefficient for the effect of birth weight on systolic blood pressure was negative,24 indicating that twin infants with lower birth weights had higher blood pressure at age 9. Heights and weights of twins at 9 and 18 years of age are not given24 but are likely to have been lower than those of singleton births. Height is established to be positively correlated with blood pressure, so it is not surprising that twins had a lower blood pressure at 9 years of age. The study by Williams and Poulton24 does not disprove the Barker hypothesis and is in direct contrast to the results from 8 year old Tasmanian children, in which low birth weight was associated with high blood pressure.25 Furthermore, when twins are studied as adults, blood pressures tend to increase as birth weight declines.26 Importantly, both these studies found that the association between blood pressure and birth weight remained in monozygotic twins, indicating that genetic factors are unlikely to be responsible for the association.25 ,26
Twins may not be a very useful model for examining the Barker hypothesis as they can vary in many ways from singletons. For example, one fetus may be aware of nutritional or endocrine signals from early pregnancy and adapt accordingly,29 whereas a singleton fetus is not subjected to such stimuli, and “spontaneous” growth restriction can occur in late gestation without any obvious clues beforehand.30 Limited epidemiological data show that twins do not have a higher mortality rate up to 6 years of age.31 Furthermore, the extent to which twin pregnancies are actually placentally and nutritionally restricted remains to be established, which is a primary criterion with respect to validation, or otherwise, of the Barker hypothesis. The relevance of the model may also depend on whether the twins are monochorionic or dichorionic. Clinically significant twin to twin transfusion syndrome affects 10% of monochorionic twins, and presumably milder forms in a greater proportion, and there is increasing evidence that intrauterine growth retardation in one twin results from poor development of the placental body supplying the smaller fetus.6 Given that angiogenesis starts at 20 days gestation in human pregnancy, intrauterine growth retardation in monochorionic twins may have little relevance to dietary nutritional deficiencies later in pregnancy.
Differences in placental and fetal development between singleton and twin pregnancies as illustrated by animal models
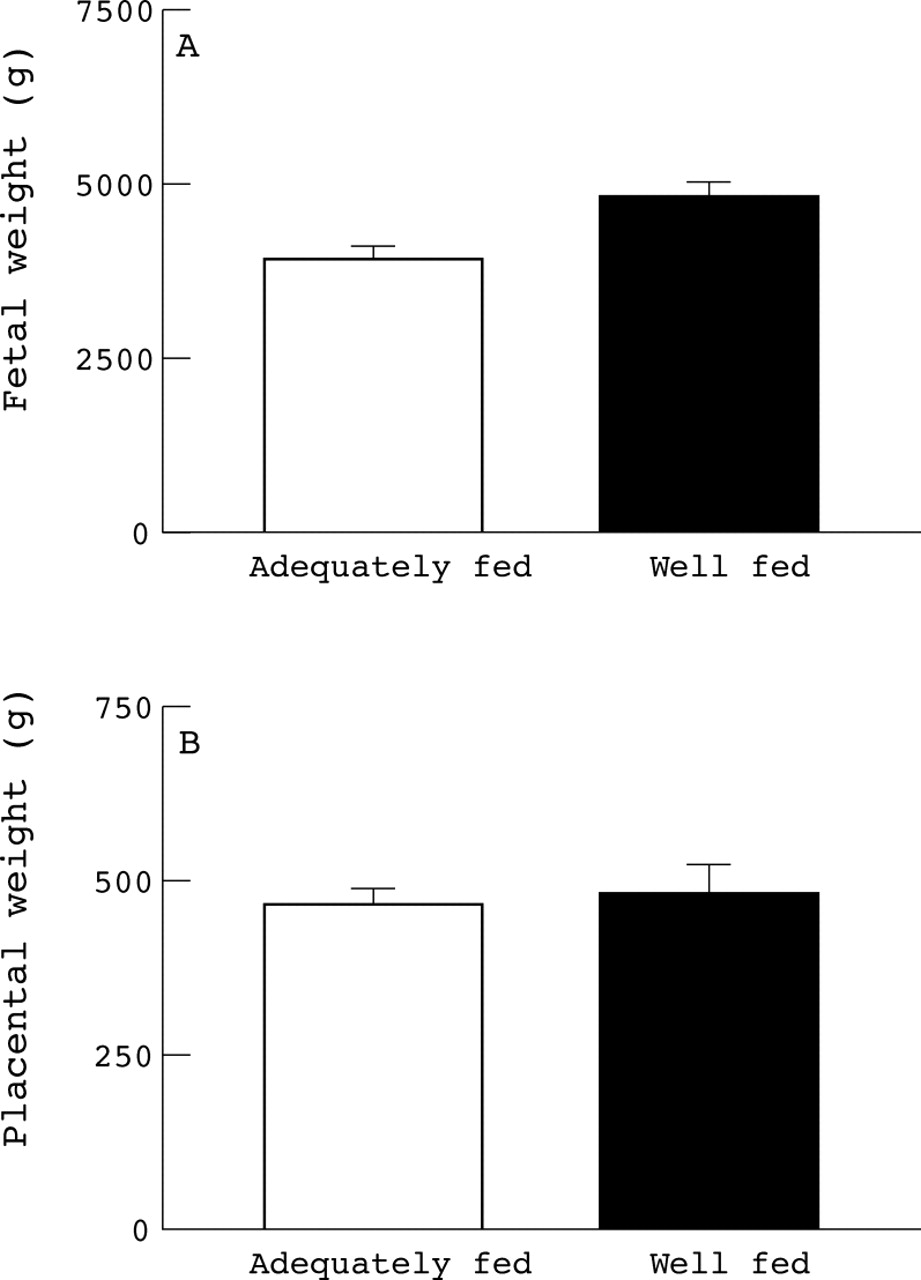
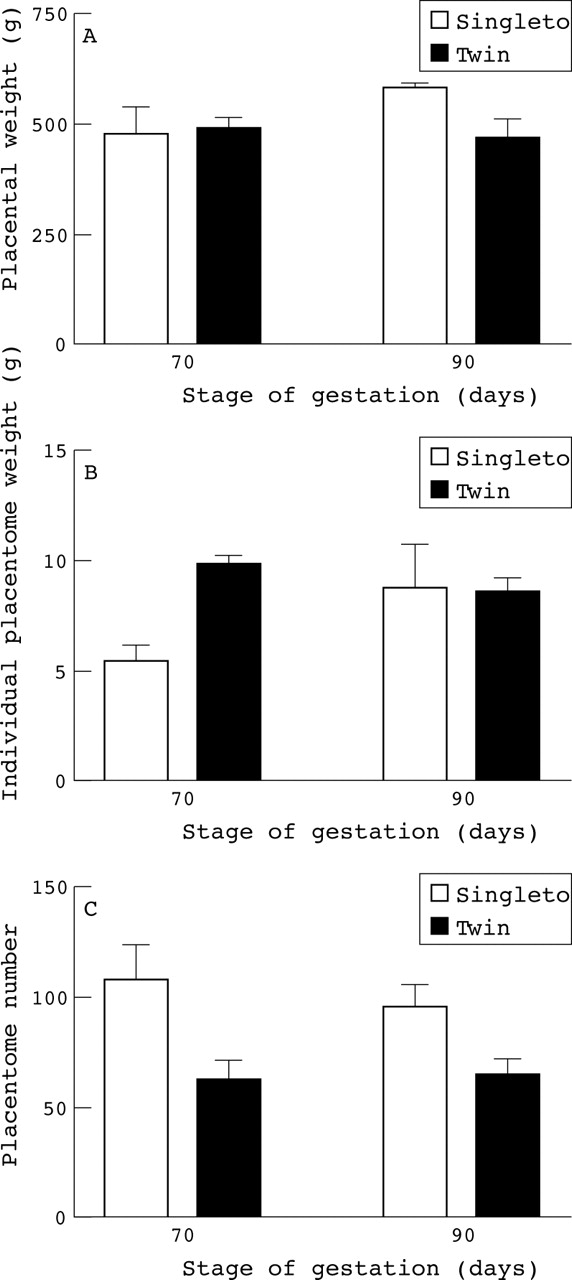
It is only possible to compare the ontogeny of placental growth with respect to fetal number using animal models. Fetal and placental development is very different between multiple and singleton pregnancies in sheep.32 At term there is a higher fetal to placental weight ratio in twin pregnancies than in singletons, although it must be noted that because ofdifferences in placentation between sheep and humans such findings can not be extrapolated directly between species. The sheep has a cotyledonary placenta composed of approximately 80 individual placentomes rather than the single disc shaped placenta of the human fetus, with the number of placentomes recruited being fixed in early pregnancy.33 At 70 days gestation there was no difference in individual total fetal weight (singletons, 150.8 ± 11.2 g (n = 4); twins, 164.0 ± 45.0 g (n = 6)) or individual placental weight between singleton and twin pregnancies (fig 4). Although each twin had fewer placentomes, mean placentome weight was substantially heavier at 70 days gestation. By 90 days gestation the individual fetal weights (singletons, 805.7 ± 30.2 g (n = 4); twins, 500.9 ± 67.4 g (n = 6) g, p < 0.05) and total placental weights per fetus for twins were significantly lower than for singletons (fig 4). These findings indicate that although the number of placentomes recruited per pregnant sheep is similar between singleton and twins, each twin has nearly half the number of placentomes. However, until mid gestation each twin has the same sized placenta as a singleton, thereby indicating that the growth of each placentome is substantially enhanced in twin pregnancies.34 It is possible that placental growth is similarly altered in twin pregnancies of humans and until one is certain of the precise differences of the nutritional and endocrine environment in which twin as compared with singleton fetuses grow and develop, one cannot be comfortable with using the birth weight and juvenile health characteristics of twins as a model for the general population.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Effect of fetal number on individual placentome weight and number in sheep at mid gestation with term being 147–8 days. (A) Total placental weight per fetus. (B) Mean placentome weight. (C) Mean placentome number. Values are means with their standard errors and were adapted from Mostyn et al.34
Conclusions
Experimental models using significant and precisely defined manipulations show that maternal nutrition does have a significant influence on placental and fetal weight and body shape, and that these effects may alter depending on the stage of gestation at which the nutritional challenge occurs. The concept that nutritional compromise at specific stages of placental, fetal, or postnatal development may directly contribute to adult disease remains an important area of debate. A study in an adequately nourished Western population relying on subjective food diaries at two discrete time points may not be the most sensitive test of this important hypothesis. Even if the effects are too small to detect by such a crude approach, they may still be important in population terms as “actions that greatly benefit the population at large may bring only small benefits for individual people”.35 Small effects may also be important in understanding mechanisms of programming in fetal life and genesis of disease in later life. Twins, which account for less than 1% of all pregnancies may not be an appropriate model for studying these hypotheses in the majority singleton population.
In summary, the hypothesis that placentofetal development has a direct influence on adult disease remains to be tested with appropriate experimental models. Relating nutrient intake at one brief time point in pregnancy to outcome at birth has a large potential to produce confounding results. Maternal nutrient intake in early to mid gestation may have a much greater influence on placental rather than fetal growth and changes in nutrition between early to mid and early to late gestation can alter both placental weight and fetal size, without any affect on birth weight. Twins studies for investigating fetal origins of adult disease in singletons are flawed owing to the profound differences in both placental and fetal development between singletons and twins. The extent to which maternal nutrition determines placental and fetal weights, as well as size at birth, remains to be fully determined, and it is only by the combination of appropriate animal models with more extensive population based studies that progress will be made.