Article Text

Abstract
Objective α-Haemolysin (HlyA) influences host cell ionic homeostasis and causes concentration-dependent cell lysis. As a consequence, HlyA-producing Escherichia coli is capable of inducing ‘focal leaks’ in colon epithelia, through which bacteria and antigens translocate. This study addressed the role of HlyA as a virulence factor in the pathogenesis of colitis according to the ‘leaky gut’ concept.
Design To study the action of HlyA in the colon, we performed oral administration of HlyA-expressing E coli-536 and its isogenic α-haemolysin-deficient mutant (HDM) in three mouse models: wild type, interleukin-10 knockout mice (IL-10−/−) and monoassociated mice. Electrophysiological properties of the colonised colon were characterised in Ussing experiments. Inflammation scores were evaluated and focal leaks in the colon were assessed by confocal laser-scanning microscopy. HlyA quantity in human colon biopsies was measured by quantitative PCR.
Results All three experimental mouse models infected with HlyA-producing E coli-536 showed an increase in focal leak area compared with HDM. This was associated with a decrease in transepithelial electrical resistance and an increase in macromolecule uptake. As a consequence, inflammatory activity index was increased to a higher degree in inflammation-prone mice. Mucosal samples from human colon were E coli HlyA-positive in 19 of 22 patients with ulcerative colitis, 9 of 9 patients with Crohn's disease and 9 of 12 healthy controls. Moreover, focal leaks were found together with 10-fold increased levels of HlyA in active ulcerative colitis.
Conclusions E coli HlyA impairs intestinal barrier function via focal leak induction in the epithelium, thereby intensifying antigen uptake and triggering intestinal inflammation in vulnerable mouse models. Therefore, HlyA-expressing E coli strains should be considered as potential cofactors in the pathogenesis of intestinal inflammation.
- Epithelial Barrier
- Epithelial Permeability
- Bacterial Pathogenesis
- Bacterial Enterotoxins
- Ulcerative Colitis
Statistics from Altmetric.com
- Epithelial Barrier
- Epithelial Permeability
- Bacterial Pathogenesis
- Bacterial Enterotoxins
- Ulcerative Colitis
Significance of this study
What is already known about this subject?
-
Proteobacteria, in particular adherent-invasive Escherichia coli, have been associated with the pathogenesis of IBD. In active ulcerative colitis (UC), the microbiota of the large intestine changes towards higher bacterial numbers of Enterobacteriaceae with E coli as main resident.
-
α-Haemolysin-carrying E coli (eg, uropathogenic E coli; UPEC) are frequently found in the faeces of humans and livestock animals. Patients with active UC develop mucosal microlesions and erosions in the colon.
What are the new findings?
-
A novel mechanism of bacterial pathology which initiates lesions in the colon mucosa called ‘focal leaks’ was identified in vivo in three different mouse models.
-
Induction of ‘focal leaks’ perturbed epithelial barrier function and increased the influx of high molecular weight antigens.
-
Colonic mucosal inflammation was accelerated by α-haemolysin-carrying E coli in inflammatory mouse models.
-
In contrast to earlier observations, the prevalence of α-haemolysin-positive E coli in human colon was high, detectable by UPEC-hlyA-specific PCR. Haemolysin-carrying E coli in human mucosa from patients with active UC were more abundant than in controls.
How might it impact on clinical practice in the foreseeable future?
-
Bacteria with the ability to impair mucosal barrier function, as for example, E coli carrying haemolysins, may become increasingly important in the epidemiology of IBD.
-
New therapeutic concepts like bacteriotherapy (eg, administration of E coli Nissle) or vaccination against UPEC could be considered for forthcoming clinical trials in IBD.
Introduction
Within the gut, most strains of Escherichia coli are regarded as commensals, some (eg, E coli Nissle 1917) even possessing probiotic properties. However, some strains (eg, enteropathogenic E coli (EPEC) or uropathogenic E coli (UPEC)) possess virulence factors which cause intestinal and extra-intestinal diseases. Furthermore, other Enterobacteriaceae, such as Salmonella and Campylobacter, may have a role in the pathogenesis of inflammatory bowel disease (IBD).1 ,2 Genome-wide association studies suggest a genetic basis to IBD susceptibility, which may involve changes in intestinal epithelial barrier function and the intestine's innate immune defence mechanisms.3 ,4 In healthy individuals, the spectrum of intestinal microbiota is relatively stable over time. However, a decrease in microbial diversity and a quantitative shift in the balance between different bacterial strains occurs, especially in active ulcerative colitis (UC), in which increases in enterobacteria (particular E coli), have been observed.5–7 Furthermore, E coli of the B2 phylogroup frequently reside in the intestinal mucosa of patients with IBD,8 ,9 and while these B2 strains are linked to urinary tract infections,10 they could contribute to disruption of colonic epithelial barrier function.
Intestinal epithelial barrier dysfunction may reflect tight junction (TJ) defects, an increase in apoptosis, the appearance of ‘focal leaks’ or erosions, or a combination of these factors. Barrier defects such as these increase the permeability of the mucosa to water and small solutes, their resulting ‘leak’ fluxes into the lumen being one of the pathogenetic mechanisms of diarrhoea in IBD. Increased permeability to small and large molecules, including potentially harmful antigens capable of eliciting immune responses, occurs in IBD.11 ,12 Thus, impaired barrier function may be cause and consequence of intestinal inflammation: in UC, apoptotic foci and erosions are early features of colonic inflammation, whereas in Crohn's disease (CD), erosions/ulcers are generally absent early in the inflammatory process.13
Luminal antigens are also absorbed transcellularly via endocytosis, which may be influenced by proinflammatory cytokines and/or bacterial virulence factors. E coli virulence factors such as fimbriae, enterotoxins and serine protease autotransporters (SPATES, eg, Sat) have been implicated in intestinal disorders.14 ,15 By contrast, the role of E coli α-haemolysin in the gut is still unclear, despite being well characterised in other respects.16 Haemolysins are pore-forming toxins (PFTs), E coli α-haemolysin (HlyA) belonging to the β-barrel-PFT (β-PFT) group. It forms small cation-permeable channels in the host cell membrane with an inner diameter of approximately 10 Å. Haemolytic bacteria have been shown to induce intestinal malabsorption,17 secretion18 and epithelial lesions via necrosis and/or apoptosis.19 We previously showed that HlyA induced ‘focal leaks’ in colonic HT-29/B6 cells, within which E coli accumulated before undergoing translocation.20 HlyA-containing supernatants also delayed epithelial restitution after induction of single-cell lesions in mouse colon epithelium.21 Thus, the aim of the present study was to determine the effect of E coli HlyA-induced impairment of colonic epithelial barrier function on antigen translocation and colonic mucosal inflammation.
Methods
Bacterial culture and intestinal colonisation model
HlyA-producing E coli-536 and E coli-536 haemolysin-deficient mutant (HDM) (536ΔhlyI_ΔhlyII::cat)22were cultured in Luria-Bertani broth at 37°C overnight and grown to log phase. For infection either E coli-536 or E coli-536 HDM was diluted to 106 colony-forming unit (CFU)/100 µL and administered via oral gavage to mice (200 µL) for intestinal colonisation according to previous protocols by pre-conditioning with streptomycin via drinking water to reduce the coliform microbiota.23 Female C57BL/6J wild type (WT) mice or B6.129P2-IL10<tm1Cgn>J interleukin-10 knockout mice (IL-10−/−) (Jackson Laboratories, Bar Harbor, Maine, USA) were used for colonisation. Furthermore, germfree C3H/ORL WT mice (Charles River, Sulzfeld, Germany) were inoculated for intestinal monoassociation with a lower infectious dose (105 E coli/100 µL). Mice were kept under germ-free conditions and were sacrificed 1 week after infection (approval number G0279/08). The intestine was removed and directly used for electrophysiological measurements. Further samples were fixed in parallel in paraformaldehyde or formalin for histological staining. Faeces were plated in different dilutions on selective agar for CFU counting. Faeces were also assessed for consistency and occult blood using hemoccult (CARE Diagnostica, Moellersdorf, Austria). Body weight of each mouse was monitored. Clinical colitis score was assessed as described previously.24
Histopathology
The tissues were fixed in a 4% formalin solution for 2 h, paraffin embedded and cut in serial sections (4 μm) for H&E staining. The pathologists were blinded to the assessment of the principal investigator. Epithelial changes were graded as described previously with a scoring range between 0 (healthy) and 12 (inflamed).25
Transepithelial electrical resistance, impedance spectroscopy and molecule marker fluxes
Mouse colon samples were mounted into Ussing chambers and total transmural electrical resistance was recorded by a computerised automatic clamp device (Fiebig-Hard&Software, Berlin, Germany). Resistance values were corrected by subtracting resistance of the bathing solution between the voltage-sensing electrodes. The total transepithelial resistance (Rt, TER) of the intestinal barrier consists of two components arranged in series, epithelial (Repi) and subepithelial resistance (Rsub). By means of impedance spectroscopy it is possible to distinguish between Repi and Rsub as previously described.26 Concurrently, unidirectional flux measurements were performed from mucosa to serosa under short-circuit conditions. 100 µM fluorescein or 18 µM HRP (Sigma-Aldrich, St Louis, Missouri, USA) was added to the mucosal side of the tissue. At specific intervals, samples were taken from the basolateral hemichamber. Fluorescence or enzymatic activity was measured in a spectrofluorimeter (Tecan, Maennedorf, Switzerland) and permeability was calculated from flux over concentration difference.
Epithelial apoptosis
Immediately after sacrifice of animals, tissues were fixed with 2% paraformaldehyde, embedded in Tissue-Tek O.C.T. medium (Sakura, Alphen, The Netherlands) for cryosectioning and cut into 5 µm cross sections, then permeabilised with 0.2% Triton X-100 and stained with rabbit-anti-cleaved-caspase-3 (1:400, Cell Signalling Technology Inc, Danvers, Massachusetts, USA) and mouse-anti-cytokeratin-20 antibodies (1:100, Invitrogen, Karlsruhe, Germany). Apoptotic cells were counted microscopically.
Visualisation of E coli HlyA and epithelial integrity by immunofluorescence
Colon tissues were fixed with 2% paraformaldehyde for 3 h without rinsing the luminal content to fix the bacteria's localisation, followed by quenching the protein linking up with 25 mM glycine. The tissue was stained as whole tissue mount without cutting as recently described.27 Briefly, the fixed tissue was permeabilised with 1% TritonX-100 in phosphate-buffered saline (PBS) for 2 h at 37°C, and subsequently incubated in blocking solution (10% goat serum, 1% bovine serum albumin, 0.8% TritonX-100 in PBS) for 3 h at room temperature. Incubation with the primary antibody in blocking solution was performed over night at 4°C. The tissue was washed with blocking solution four times with a prolonged washing time of 1 h each, then incubated over night at 4°C with a secondary IgG antibody and washed again four times. Staining with DAPI and Phalloidin-AlexaFluor647N was performed at room temperature for ½ h (each 1:1000). The following antibodies were used: anti-HlyA, anti-E coli-O (1:200 provided by Lothar Beutin), anti-occludin (1:100), anti-E-cadherin (1:250), AlexaFluor-488 goat-anti-mouse, AlexaFluor-488 goat-anti-rabbit, AlexaFluor-594 goat-anti-mouse or AlexaFluor-594 goat-anti-rabbit IgG (1:500; Invitrogen). Antibody sensitivity was evaluated in E coli-536-infected HT-29/B6 cell culture. HT-29/B6 monolayers were grown on permeable polycarbonate filters in antibiotic-free RPMI 1640 medium as described previously.20 Stained tissues were mounted using ProTaqs MountFlour (Biocyc, Luckenwalde, Germany). Stainings were visualised by confocal laser-scanning microscopy (C-LSM, Zeiss LSM510, Jena, Germany) with a 40× water-immersion objective (2 mm working distance for deep optical XY-plain sections in Z-stacks).
Measurement of hlyA in human colon biopsies
Biopsy specimens were taken from sigmoid colon from patients undergoing routine endoscopy (approval number EA4/098/09). In patients with active inflammation the tissue was taken from minor inflamed areas. DNA extraction was performed according to the manufacturer's instruction by bead beating (ZR Fecal DNA MiniPrep, Zymo Research, Irvine, California, USA). Quantitative real-time PCR (qPCR) was performed using SYBR-green with an ABI 7900HT PCR device according to manufacturer's instructions (Applied Biosystems, Mannheim, Germany) with EHEC-hlyA type-specific primers8 or UPEC-hlyA type: HlyA536_for: 5′-TTCTGCTGTGACACTGGCAA-3′, HlyA536_rev:5′-TAACAGCACCTACCAGTGCG-3′ and the values were referred to a standard curve and compared with total bacteria/E coli with primers used previously (see online supplementary tables S1–S3).28 ,29
Statistical analysis
Data are expressed as mean values±SE of the mean (SEM). Statistical analysis was performed using two-tailed Student's t test or Mann–Whitney U test for non-parametric data, as appropriate. p<0.05 was considered statistically significant.
A detailed description of methods can be found in the online supplements.
Results
Intestinal disorder in challenged animals
Oral infection of mice with 106 E coli-536 or E coli-536 HDM revealed colonisation of the gastrointestinal tract between 106 and 108 CFU/g faeces 48 h after infection. All successfully colonised mice were assessed for their outcome (diarrhoea and colitis activity). Infected mice revealed no or only moderate symptoms, assessed by body weight, faecal consistence and occult blood. Mice infected with the α-haemolysin-expressing E coli-536 showed increased colitis activity compared with controls infected with the HDM strain. As already reported, the E coli-536 colonised WT mice showed no or only moderate clinical symptoms,23 whereas an increase in colitis score was measurable in the IL-10−/− colitis model and in the monoassociated (MA) mice compared with the HDM-infected animals (figure 1A).

Colitis activity. (A) Colitis scores measured in mice after oral infection with Escherichia coli-536, compared with α-haemolysin (HlyA)-deficient E coli 536 haemolysin-deficient mutant (HDM) strain, either in wild type (WT), interleukin-10 knockout mice (IL−10−/−) colitis model or monoassociated (MA) WT mice. Dots were single values; bars represent mean values ranging from 0 (healthy) to 4 (maximal activity of colitis). **p<0.01,***p<0.001, n.s., not significant; Student's t test. One of seven HDM-infected IL-10 mice was not colonised and excluded from analysis. (B) Pathohistology; H&E stainings of colon mucosa of 536-colonised WT, IL-10−/− or MA mice. For comparison, healthy mucosa without lymphocyte infiltration: on the left side HDM-colonised WT or MA mouse colon. Depicted is one representative image of each group in quadruplicate. Ctrl, control.
Histopathology of the large intestine colonised with E coli-536
The clinical symptoms had their correlate in colon histology. The mucosa of E coli-536-challenged mice showed mild changes with sporadic lesions, focal loss of enterocytes, enlarged lymph follicles and slightly increased lymphocyte infiltration into the lamina propria. The histological appearance of colonised WT mice was almost normal, whereas the germfree WT mice exhibited moderate histological changes when infected with E coli-536 for monoassociation (figure 1B). Most importantly, the IL-10−/− mice showed increased histopathological responses to E coli-536 in comparison to HDM. Changes were not as severe as in active colitis, but reflect an initial/early phase of inflammation. The 536-treated IL-10−/− mice exhibited a higher colitis score of 1.8±0.4 versus 0.6±0.2 in controls (p<0.05, n=5, animals between 3 and 8 days post infectionem). The overall epithelium was intact, fulfilling the prerequisite for subsequent electrophysiological measurements of epithelial barrier function.
Electrophysiological observations on the colon of challenged mice
Barrier function of the mucosal epithelium was assessed in Ussing experiments. Impedance spectroscopy measurements revealed a decrease in overall transepithelial electrical resistance (Rt) in the colon colonised with HlyA-harbouring E coli-536 compared with mice colonised with strain 536-HDM (figure 2). This loss of electrical resistance of the colon specimens could be due to a drop in epithelial (Repi) and in subepithelial (Rsub) resistance. In the case of the MA mouse model, the drop in Rt could be assigned to a drop in Repi, while Rsub remained unaffected. In WT and IL-10−/− mice, Rt was also decreased and Rsub remained almost unaltered but the concomitant reduction in Repi just failed to reach statistical significance.

Electrical resistance of mouse colon measured by impedance spectroscopy. Overall transmural electrical resistance (Rt, dark grey bars) consists of epithelial resistance (Repi, light grey bars) and subepithelial resistance (Rsub, bright bars). The left panel shows the electrical resistance of wild type (WT) mice infected either with the haemolysin-deficient mutant (HDM) of Escherichia coli as control or with the α-haemolysin (HlyA)-carrying E coli-536 (n=7 and 8). The middle panel shows the results obtained from interleukin-10 knockout mice (n=6 and 8) and the right panel measurements of monoassociated (MA) WT mice (n=5 and 5) colonised in the same manner by the E coli strains. *p<0.05 (Student's t test).
In parallel, macromolecule permeability measurements with 44 kDa horseradish peroxidase (HRP) were performed and revealed an increase in HRP fluxes in all 536 infected mice compared with HDM controls (figure 3). The already initially increased flux in the IL-10−/− mice can be interpreted as the pre-existing pathogenic mechanism for antigen/HRP antigen uptake prior to colitis. Fluorescein (332 Da) was used as an additional marker, which in the case of unaltered macromolecule flux and thus the absence of gross lesions would have been a parameter for disturbed TJs. A slight increase in epithelial fluorescein permeability was observed in WT and MA mice (536-WT mice with 1.04±0.11 vs 0.47±0.17 nmol/cm2/h1 HDM-control and 536-MA mice with 0.79±0.10 vs 0.41±0.11 nmol/cm2/h1 HDM-control; p<0.05, n=5 each). However, this was only a small increase with doubled fluorescein fluxes. In the case of the 536-infected IL-10−/− colitis model this increase to 0.91±0.21 nmol/cm2/h1 (n=6) even failed to reach statistical significance compared with HDM-control mice (0.51±0.10 nmol/cm2/h, n=5). Thus, this small increase in fluorescein flux does not point to severe TJ defects but may be explained by the appearance of focal leaks.

Antigen permeability of mouse colon measured in Ussing chambers. Horseradish peroxidase (HRP) fluxes from mucosal to serosal compartment were assessed by enzymatic reactivity in the serosal compartment. *p<0.05 (three to four animals per group, Student's t -test). Crtl, control; HDM, haemolysin-deficient mutant; HlyA, α-haemolysin; IL-10–/–, interleukin 10 knockout; MA, monoassociated; WT, wild type.
Epithelial apoptosis
Investigation of epithelial cell death—specifically the search for focal areas of increased apoptotic cell ratio—was done in the colon after staining of cleaved caspase 3. We could detect only very few sporadic apoptotic foci in IL-10−/− and MA HlyA mice, while single apoptotic events were present throughout all specimens (figure 4A). However, the apoptotic ratio in WT, IL-10−/− and MA mice infected with E coli-536 remained statistically unchanged compared with control HDM mice (figure 4B) and also no difference was obtained between the different models.

Epithelial apoptosis. (A) Epithelial cell apoptosis in colonic tissue of Escherichia coli-536 infected mice detected with anti-caspase-3 immunofluorescence microscopy 5 days after infection. Tissues from infected animals were immediately fixed after sacrification. Single apoptotic events were located evenly throughout the colonic epithelium, whereas apoptotic fociwere detected only sporadically. Depicted is one representative image of at least three animals. (B) Quantification of cleaved caspase-3-stained apoptotic events (apoptotic rate) was performed in wild type (WT) (n=3), interleukin 10 knockout (IL-10−/−) (n=4), monoassociated (MA) (n=3) mice colon tissue colonised with E coli 536 (HlyA) or the haemolysin-deficient mutant (HDM) control strain (given as % apoptotic cells in a low-power field). Four different serial tissue cross sections of each animal were counted with at least 1000 DAPI and cytokeratin-20-positive epithelial cells (n-values represent the number of animals in the analysis). DAPI, 4′-6-diamidino-2-phenylindole dihydrochloride, colours nuclei blue; n.s., not significantly different in Student's t test.
Confocal laser-scanning microscopy revealed lesions in the presence of HlyA-positive E coli
To detect possible focal leaks as observed in vitro earlier,20 we searched microscopically for lesions and bacterial signals in immunofluorescence (IF) stainings of unsectioned mouse colon. Lesions and/or erosions in the colonic mucosa of E coli-536-colonised mice were observed in IF stainings with co-localisation of invaded E coli (figure 5A). The mucosal lesions induced by E coli-536 showed the same shape and size as observed earlier in rat colon and HT-29/B6 cell cultures 20 (figure 5B) with a funnel-shaped notch including bacterial signals inside (figure 5C and see online supplemental movie).
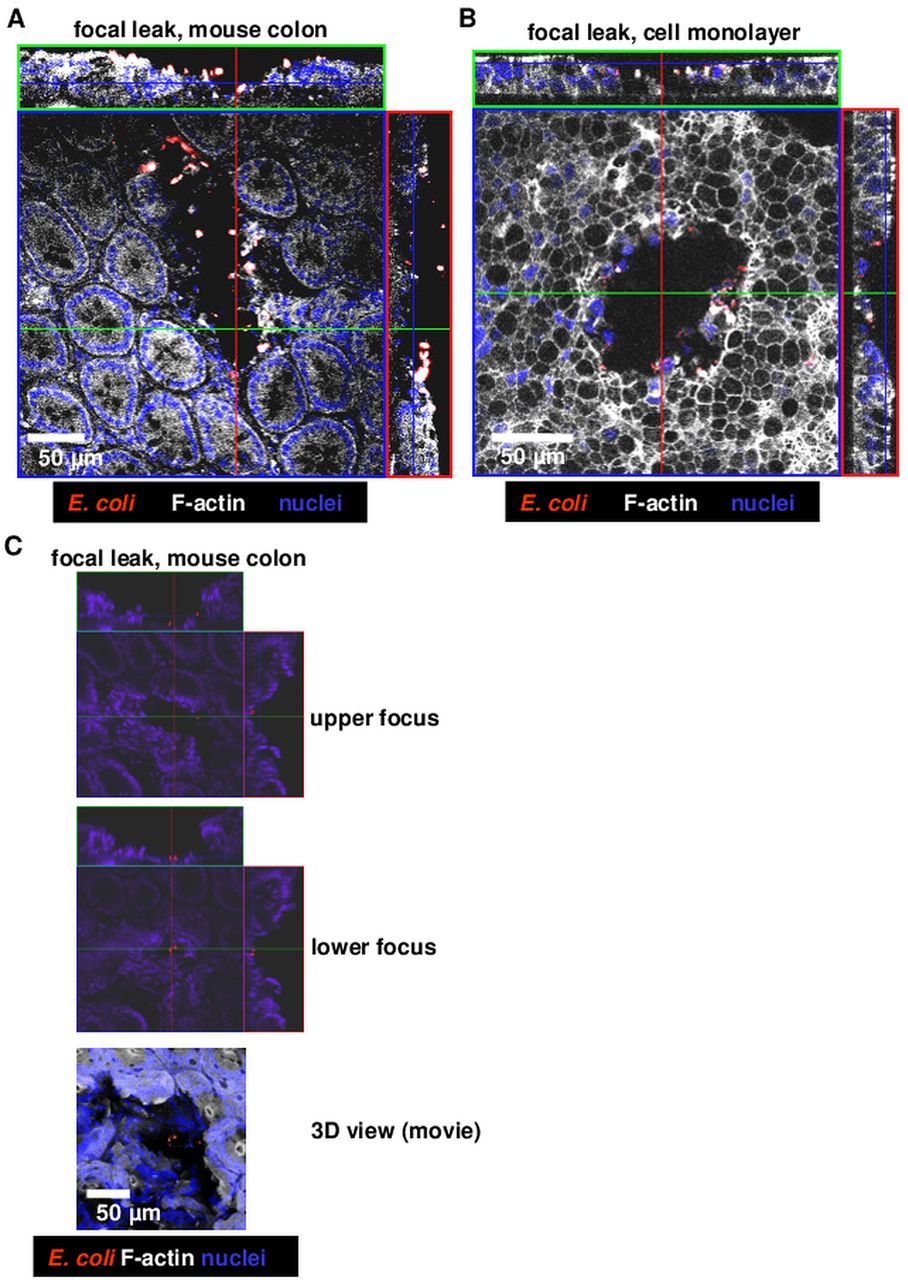
Confocal laser-scanning microscopy of immunfluorescence stainings of Escherichia coli α-haemolysin (HlyA)-induced lesions. Whole mount staining of mucosae from challenged mice or epithelial cell monolayers (without sectioning). (A) Focal leak in E coli-536-colonised wild type mouse colon, (B) focal leak in a human colon epithelial cell monolayer HT-29/B6, (C) smaller focal leak with concomitant bacterial invasion in mouse colon, see also online supplemental movie for 360° 3D view.
Size estimation of focal leaks
For demonstrating bacterial IF signals MA mice were studied in more detail (figure 6A). Focal leaks and bacterial signals could be detected in E coli-536 but not in E coli-536-HDM-colonised mucosa. For quantification, focal leaks were counted microscopically, digitally marked and sized. We found one to four gross leaks with a size range of 2000–50 000 µm2 per single leak in areas of 0.049 cm2 (equalling 4 900 000 µm2; the tissue area of the Ussing experiments). An accumulated mean leak area of 54 000 µm2 was measured in the 536-MA mice, which is 1.1% of the respective tissue area. Equivalent structures to focal leaks can also be found in the 536-infected WT mice (n=7, figure 6B) but no such leaks were detected in E coli-536-HDM-colonised WT mice. However, the IL-10−/− mouse model showed a few focal leaks when colonised with HlyA-negative E coli, though not as large as observed in the E coli-536-infected IL-10−/− mouse, probably due to the already ongoing inflammatory stage (n=3, figure 6B). The quantification of leak size revealed an increase for each mouse model with E coli-536 versus HDM colonisation. A calculation of epithelial resistance of an affected epithelium (Repi_leak) with 1.1% leak area fits to the measured values in impedance spectroscopy (see online supplemental calculation). Moreover, the overall leak area of E coli-536-infected HT-29/B6 cell monolayers increased with bacterial burden (figure 6C).
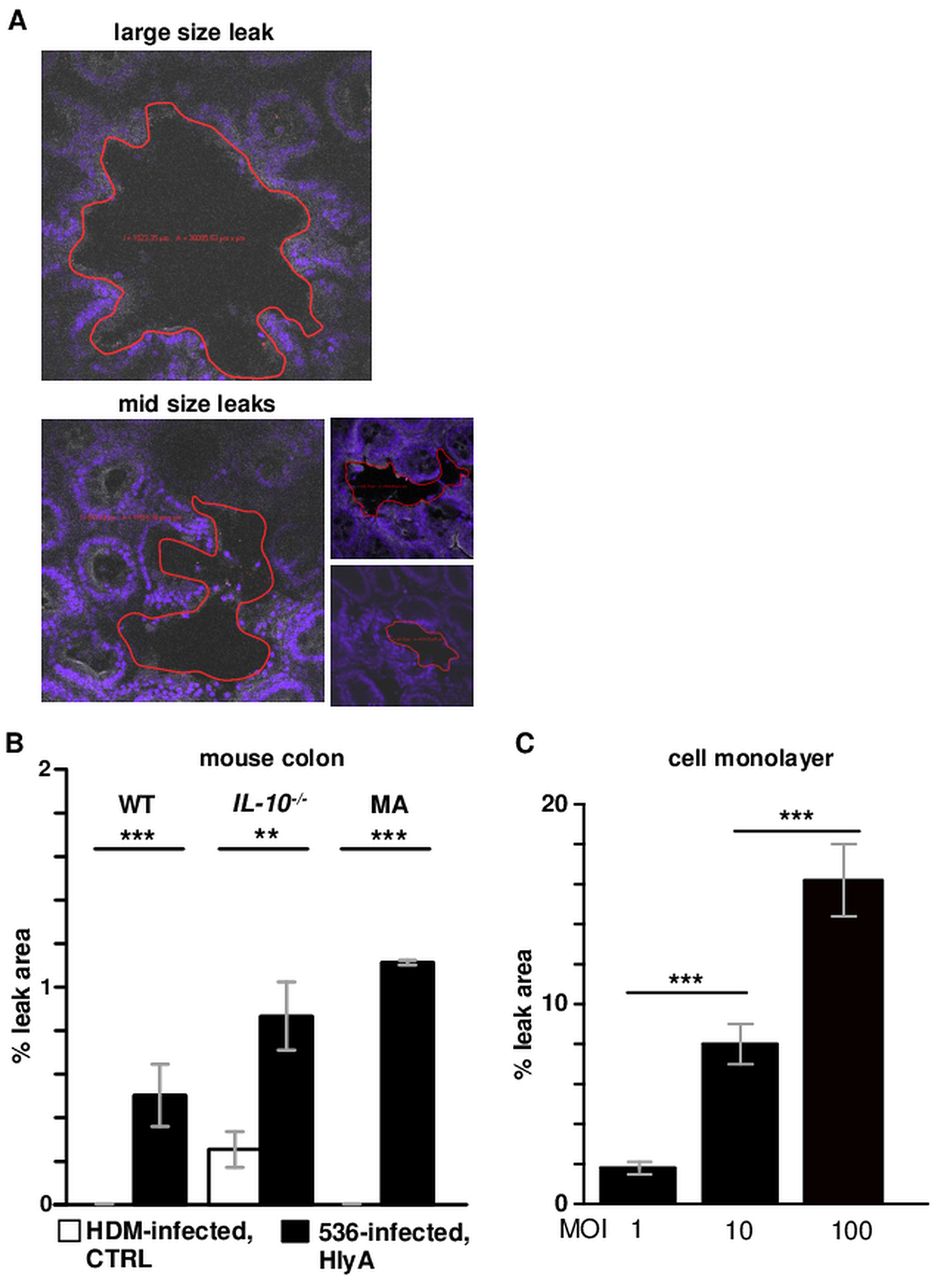
Microscopic size estimation of focal leaks. (A) Examples of large and mid sized leaks, marked (red line) and measured with Zeiss LSM Image Examiner software for focal leak area size calculation. (B) Quantitation of leak area as % of the whole investigated area in a low-power field. In whole mount immunofluorescence staining of uncut mouse colon, an observation area of approximately 0.1 cm² was screened microscopically for focal leaks. The identified leaks were digitally marked and the overall area of one to four single leaks was divided by the overall measuring area. In haemolysin-deficient mutant (HDM) treated interleukin 10 knockout (IL-10−/−) mice focal leaks were revealed, whereas in HDM-treated wild type (WT) and monoassociated (MA) mice no leaks were observed in three independent stainings of at least three animals (n=3). (C) Dose-dependent appearance of focal leaks in a human colonic HT-29/B6 epithelial cell monolayers after 3 h treatment with different multiplicity of infection (MOI) of Escherichia coli-536 (n=3 each MOI). **p<0.01,***p<0.001, n.s., not significant; Student's t-test. HlyA, α-haemolysin.
HlyA increased in human colon mucosa during active UC
As reported previously, hlyA was not detected in bacterial cultures from biopsy specimens using EHEC-specific hlyA-primers.8 ,30 However, in qPCR measurements using our primer for UPEC-specific hlyA-type and DNA extraction from biopsy samples without pre-cultivation of bacteria, we found hlyA-positive E coli in 19 of 22 patients with UC, 9 of 9 patients with active CD and 9 of 12 healthy controls (see online supplementary tables S1–S3). While prevalence between healthy controls and patients with IBD was not different, there was a more than 10-fold increase in hlyA level in mucosal biopsies of active UC compared with healthy controls (p<0.05, figure 7A). The total bacterial count was not significantly altered between the groups (7110±3620 copy numbers/ng DNA in active UC vs 4150±3700 copy numbers/ng DNA in healthy controls) but showed a tendency towards an increase in Enterobacteriacea as indicated by Eco primer (figure 7A). The increase in bacterial numbers harbouring E coli HlyA seemed to be specific for UC, since high concentrations of HlyA were neither found in colon samples from active CD nor in healthy individuals (figure 7B). Moreover, it was possible to find focal leaks in human sigmoid colon in patients with UC by confocal endomicroscopy with biopsy sampling and subsequent IF staining and C-LSM of the tracked biopsies (figure 7C, see online supplementary figure S1). In the colon biopsies of non-inflamed regions from patients with UC small lesions could be pictured which showed HlyA-positive signals inside.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
α-Haemolysin (HlyA) in human colon. (A) Haemolysin measurements in human colon biopsies by quantitative PCR. Number of total bacteria (total), of Escherichia coli (Eco) or of HlyA 536-positive E coli per PCR specimen (1 ng DNA, equalling approx. 10 µg mucosal biopsy) of 12 healthy controls compared with 11 patients with ulcerative colitis with active colitis. *p<0.05 in Mann–Whitney U test. (B) Percentage of patients with concentrations of HlyA-positive mucosal E coli. PCR copy numbers as indicated per colon biopsy specimen (1 ng DNA) from 12 healthy controls, 9 patients with active CD, 11 patients with UC in remission and 11 patients with active UC. (C) Focal leak with HlyA-positive signals in human colon mucosa. Detail of a focal leak at the edge of a crypt. Confocal laser-scanning microscopy on whole mount biopsy specimens after immunfluorescence staining obtained from a patient with inactive UC as one representative image in triplicate. Intact endogenous control area without HlyA signal and a focal leak with appearance of HlyA (red), while E-cadherin (green) and F-actin (white) are absent at the edge of a crypt. Nuclei are coloured blue. The insert on the right side shows a detail with F-actin staining and HlyA signals at and in the epithelium. See online supplementary figure S1 for the overview of the affected region within a non-inflamed area and the workflow.
Discussion
Induction of focal leaks by E coli HlyA
E coli HlyA-induced focal leaks—originally discovered in HT-29/B6 monolayers20—were also detected in three in vivo mouse models. Generally, the UPEC strain 536 used in our present study has a different genetic constitution compared with attaching and effacing (A/E) E coli, which produce typical lesions with pedestal actin formation, mediated by effector proteins like intimin or EspF.31 In contrast to a typical EHEC, E coli-536 lacks the pathogenicity island locus of enterocyte effacement for A/E lesions. Thus, we proposed and confirmed an alternative pathomechanism for HlyA in initiating lesions in the intestinal mucosa.
Cellular response mechanisms to HlyA
A/E-typical pedestal actin formation was not seen in microscopic images of E coli-536-colonised epithelium. However, HlyA is known to create channels in the host cell membrane that promote potassium efflux, affect Akt/protein kinase B activity 32 and enable calcium influx into the host cell with various consequences resulting from calcium-dependent cell signalling. Regardless of the contribution of host cell calcium channels, a cation-selective pore like HlyA itself can mediate a calcium influx which may lead to cytoskeleton alterations or cell lysis. Rapid calcium oscillations have been described in renal epithelial cells in response to UPEC HlyA.16 The calcium-dependent response of enterocytes to β-PFT haemolysins in sublytic concentrations shows different cellular characteristics, including impaired epithelial restitution, cytoskeletal and TJ disturbance as reported for aerolysin (AerA) from Aeromonas hydrophila.33 In contrast, characteristic focal leaks as induced by HlyA were not found in AerA-treated intestinal epithelial cells.33
The various pathogenic impacts of β-PFTs could be explained by differences in expression or the site of infection, and by toxin polymorphisms. Even strain-specific differences in genomic sequences may have an impact on their different modes of pathogenic action (eg, in E coli HlyA only ∼70% sequence identity between EHEC and UPEC HlyA types exists). However, the role of calcium influx through the HlyA pore in epithelial cells, resulting in signalling events which possibly cause focal leak formation, needs further investigations. Hypothetical modes of focal leak creation include epithelial damage (eg, via cell death mechanisms such as necrosis or autophagia, while apoptosis could be excluded), and/or the inhibition of cellular restitution of small lesions.
Disruption of the epithelial barrier by HlyA
Epithelial barrier function of the colon was seriously compromised by HlyA. This was less evident from measurements of electrical resistance, which was only slightly affected in HlyA-positive E coli 536-infected mice. However, this was clearly evident from Ussing-chamber flux measurements with larger molecules. We decided to use HRP as antigen influx marker because it has two advantages: it is a suitable marker for passage through leaks as it cannot diffuse paracellularly through the TJ; and it can be measured enzymatically which guarantees that the molecule remains uncleaved and avoids misinterpretation from released labels. Thus, the increased HRP flux measured in this study can be explained by the appearance of focal leaks in all three mouse models after E coli-536 colonisation. The size estimation of focal leaks induced by HlyA-carrying E coli-536 revealed that approximately 1% of the observed mucosa was affected, and a calculation of the increased conductivity of the leaky epithelium revealed that this focal leak size explains most of the change in epithelial resistance (see online supplemental calculation). However, we cannot exclude that other pathological effects induced by E coli-536 such as TJ changes with strand discontinuities may have an additional impact. We also excluded HRP transcytosis as an alternative mechanism of macromolecule uptake by histochemistry. No increase in endocytosis by colonocytes was seen (see online supplementary figure S2), which would have been an electrically silent process, not influencing transepithelial resistance measurements. TJ changes could only be observed around focal leaks (see online supplementary figure S3 as eg, condensation of occludin) and global changes in strand forming TJ proteins, such as claudins 1–3, could be ruled out by Western blot densitometry (see online supplementary figure S4).
Accelerated inflammation by α-haemolysin-expressing E coli
In addition, α-haemolysin-expressing E coli enhanced inflammation in our inflammatory mouse models. As already reported, this inflammatory response was not measurable in WT mice,23 but was present in the vulnerable IL-10−/− colitis model. A difference was also seen in HlyA-carrying versus HDM-E coli groups in the MA mice in the initial stage of colitis. Such mild but significant effects resulting from the onset of colitis could also be assessed in other studies with mild inflammation in IL-10−/− mice.34
α-Haemolysin-expressing E coli (eg, UPEC) were frequently found in faeces of healthy humans and livestock animals.35 ,36 Within the healthy mucosa of the colon, no intraepithelial bacteria should be present. Only in cases of infection (eg, EHEC), or in the presence of inflammation (IBD), do high numbers of mucosa-associated or intraepithelial E coli invade the colonic mucosa, but so far without evidence of any correlation between the severity of inflammation and the bacterial count.37–40
High prevalence of hlyA, especially in UC biopsies
qPCR measurements in human biopsy specimens with UPEC hlyA-type specific primers revealed an overall high prevalence of hlyA in the colon mucosa, but higher levels in patients with active UC. This is in line with previous findings of high hlyA occurrence in human stool samples in a non-inflamed stage.35 Moreover, Martinez-Medina and colleagues9 found hlyA in about 20% of E coli isolates from the biopsies of 10 patients with CD and 12 healthy control patients with similar primers, and in another study using Chip technology for comparative genomics, E coli isolates from the biopsies of two of five patients with UC were positive for hlyA.41 Our findings could depict for the first time that HlyA is indeed usually present, but much more frequently in colonic samples of patients with active UC, and thus may play a significant role in intensifying inflammation. HlyA may play a role in initiating lesions in UC, since we could identify HlyA in IF stainings inside and around lesions of endoscopically unaffected tissues from patients with mild inflamed disease. It is conceivable that HlyA-carrying E coli have easier access to the colonic mucosa when it is inflamed.40 In addition, HlyA is frequently associated with bacterial outer membrane vesicles, which may contain a cocktail of various proteins/toxins from the bacterial periplasm, including factors contributing to facilitated access to the mucosa.
Conclusion
Our findings suggest that E coli HlyA can induce focal leaks in cell models, and in active UC and native mouse colon, and thereby increase antigen invasion as indicated by HRP flux measurements, providing an additional pathological mechanism for antigen influx consistent with the ‘leaky gut concept’. Furthermore, this study shows that extra-intestinal pathogenic E coli, when colonising the intestine in high numbers, trigger inflammation or act as inductors prior to inflammation in quiescent colitis. Thus, erosions and ulcers observed in UC could develop from increased apoptosis and/or be initiated via mediators, such as HlyA.
Selective reduction of bacteria with barrier-breaking features (eg, haemolysins) raises the possibility of new treatment options in patients with IBD. Future studies may prove that vaccination against certain Enterobacteriaceae (as in chronic cystitis) or bacteriotherapy with (for example) non-pathogenic E coli strains, are new and effective alternative therapies for IBD.
Acknowledgments
The excellent technical assistance of Detlef Sorgenfrei and Anja Fromm is gratefully acknowledged. We thank Gábor Nagy who constructed E coli-536-HDM. This work is dedicated to Professor Martin Zeitz († 26 November 2013).
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Files in this Data Supplement:
- Data supplement 1 - Online supplement
- Data supplement 2 - Online video
Footnotes
-
Contributors Conceived and designed the experiments: RB, JDS, HT. Performed the experiments: RB, ES, I-FML, SW, LJJ, TJ, CE. Acquisition of biopsy material from endoscopy: CB, ES. Analysed the data: RB, ES, I-FML, LHW, J-DS, HT, DG, CB. Important intellectual support: ES, UD, LB, LHW, DG, CE, MF, BS. Contributed reagents/materials/analysis tools: TJ, LHW, UD, LB, LJJ, DG, CB, MF, JDS. Wrote the paper: RB. Critical revision and study supervision: MF, J-DS.
-
Funding Supported by Deutsche Forschungsgemeinschaft (Schu 559/11 and SFB 852).
-
Competing interests None.
-
Patient consent Obtained.
-
Ethics approval Animal experiments were approved by the local governmental authorities under the approval number G0279/08 (LaGeSo, Landesamt für Gesundheit und Soziales, Berlin, Germany). For the use of human material this study adhered to the Declaration of Helsinki, and ethics approval for research was obtained from The Ethics Committee of the Charité Berlin (approval number EA4/098/09 and EA2/179/05). All patients who participated in the investigation signed written informed consent forms. Children were not included in the study.
-
Provenance and peer review Not commissioned; externally peer reviewed.