Article Text

Abstract
OBJECTIVES Particulate air pollution has been shown to cause adverse health effects, and the ultrafine particle component has been implicated. The aim of the present study was to investigate whether an ultrafine particle exerted its effects through transition metals or other soluble factors released from the surface of the particles.
METHODS Both in vitro and in vivo models were used to test the imflammogenicity of carbon black (CB) and ultrafine carbon black (UfCB) and the role of transition metals was investigated by treating the particles with desferrioxamine mesylate (desferal), a transition metal chelator. Rats were instilled with particles and the cell population assessed by bronchoalveolar lavage (BAL). Calcium homeostasis in macrophages was assessed with a fluorimetric technique.
RESULTS UfCB was inflammogenic compared with CB when instilled into Wistar rat lungs, an effect which could not be ameliorated by desferal treatment of the particles. Particle leachates produced no significant inflammation in vivo. In vitro experiments showed that the cytosolic calcium ion concentration in Mono Mac 6 cells was increased significantly after UfCB treatment and treatment of particles with desferal did not alter these effects. Particle leachates had no effect on cytosolic calcium ion concentration. Iron was not detected in leachates of the particles with the desferal assay, however, ng/mg of particles were detectable in citrate leachates with inductively coupled plasma-mass spectrometry (ICP-MS).
CONCLUSIONS The increased inflammogenicity of UfCB compared with CB cannot be explained by soluble transition metals released from or by accumulation of iron at the particle surface. Differences may be accounted for by increased surface area or particle number.
- ultrafine
- calcium
- transition metals
Statistics from Altmetric.com
There has been growing interest in the pulmonary response to small diameter particles with regard to their role in the adverse health effects of particulate air pollution.1 Ultrafine particles have been shown to have marked toxicity,2 and it has been hypothesised that the ultrafine component of particulate air pollution (PM10) is responsible for their adverse effects.2 3 The precise mechanisms whereby ultrafine particles exert their effects in the lung are not fully understood.2 In long term high dose inhalation studies in animals, the chronic effects produced by ultrafine particles include inflammation, increased chemokine expression, epithelial hyperplasia, pulmonary fibrosis, and lung tumours.4-8 However, these effects are a consequence of overload.9 Short term low dose non-overloaded inhalation exposure to ultrafine carbon black (UfCB) has produced mild inflammation, oxidant stress, and modulation of the coagulation system in normal rats.10
An important part of the ultrafine hypothesis is to dissect out whether the ultrafine habit is itself inflammogenic or whether ultrafine particles simply provide a large surface area for transition metals to be released. Samples of environmental particulate—such as residual oil fly ash (ROFA)11 and Provo PM10 12—have been shown to cause inflammation by mechanisms mediated by transition metals—that is, a soluble preparation of the particles contains all the proinflammatory activity. We have also shown that soluble components of PM10 that can be chelated—that is, transition metals—are the source of oxidative stress in PM10.13 To understand what the relative toxic potency of the components of PM10 are it is necessary to know the toxicity of the individual components. We have used fine carbon black (CB) and UfCB to answer the question “can ultrafine particles cause toxicity independently of soluble transition metals?” The importance of showing whether ultrafine particles cause toxicity through mechanisms other than transition metals lies in a better understanding of the total hazard associated with any PM10 sample. If ultrafine particles could be shown to have effects through other mechanisms than by transition metal then simply measuring the transition metals does not describe the potential toxicity. We therefore used UfCB as a model ultrafine particle and set out to find whether transition metals had any role in its ability to cause increased inflammation in rat lungs and proinflammatory effects in vitro.
Therefore, the aim of the present study was to elaborate the role of transition metals in the proinflammogenic effect of UfCB with both in vitro and in vivo models. In the in vivo model, particles were instilled into the lungs of rats and the number of neutrophils was assessed by bronchoalveolar lavage, as a measure of inflammation. We used the transition metal chelator desferrioxamine and also used leachates of the soluble component, which would contain any transition metals, to investigate the role of transition metals in the proinflammatory effects of UfCB. The in vitro model is one where we have previously shown a greater effect of UfCB than CB on the cytosolic calcium ion concentration.22 The Ca2+ system is an important signalling pathway for the activation of the proinflammatory transcription factor NF-κB and other proinflammatory transcription factors which are regulated through several second messengers.15 Ca2+ is stored in the endoplasmic reticulum (ER), from where it is released on stimulation of the cell. Ca2+ release from the ER leads to the activation of calcium ion influx across the plasma membrane through calcium channels. Changes in calcium ion flux within the cell have been shown with different pathogenic particles16 17 which suggests a role for calcium homeostasis in the proinflammatory effects of particles.
Materials and methods
PARTICLE CHARACTERISTICS
Fine carbon black was obtained from H Haeffner, Chepstow, UK (Huber 900) and UfCB obtained from Degussa (Printex 90). The average particle size of the two types of particle were 320 nm for CB and 14 nm for UfCB.
PREPARATION OF PARTICLE LEACHATES
The CB and UfCB were suspended in citrate phosphate buffer pH 4.6 and 7.2 at a mass concentration of 10 mg/ml. All buffers and reagents were made up in chelex treated water. The particle suspensions were sonicated for 2 minutes to aid dispersion and mixed on a rotary mixer for 24 hours at 37°C. After incubation, the suspensions were transferred to 50 ml centrifuge tubes and spun at 145g for 20 minutes. Leachates were pipetted into clean tubes and filtered through 0.22 μm filters. The Fe3+ concentration in each leachate was estimated as outlined below.
A second series of particle leachates were prepared as before in saline and pH 7.2 citrate buffer at a concentration of 2 mg/ml for use in in vivo experiments. Leachates were prepared immediately before instillation into rat lungs.
PREPARATION OF PARTICLE LEACHATES FOR ELEMENTAL ANALYSIS
Particles were suspended in saline or citrate buffer pH 7.2 at a concentration of 1 mg/ml, sonicated to disperse the particles and incubated for 24 hours at 37°C. Particle suspensions were then centrifuged at 145 g, and the supernatant was collected and filtered through 0.22 μm filters. Each filtered sample was diluted 1/50 with chelex treated water and analysed with inductively coupled plasma-mass spectrometry (ICP-MS).
DESFERRIOXAMINE TREATMENT OF PARTICLES
The CB and UfCB were suspended in either chelex treated water or 10 mM desferrioxamine mesylate (desferal, Sigma, Poole, Dorset, UK) at a particle mass concentration of 1 mg/ml and incubated in a shaking water bath for 2 hours at 37°C. After incubation, samples were centrifuged at 145 g, the supernatant was removed, and the particles washed twice with saline. Finally the particles were resuspended in saline at a concentration of 1 mg/ml.
PARTICLE AND PARTICLE LEACHATE INSTILLATION
Female Sprague Dawley rats about 4 months old were used in all studies. The animals were anaesthetised with halothane, cannulated with a laryngoscope to expose the trachea and 0.5 ml of a 1 mg/ml particle suspension contained in saline instilled into the lungs. A separate group of animals received 0.5 ml appropriate particle leachate. All animals were conscious within minutes of this procedure and suffered no ill effects. Treated animals were killed after 24 hours.
BRONCHOALVEOLAR LAVAGE
Rats were killed with a single intraperitoneal injection of pentobarbital, and the lungs were cannulated, removed, and lavaged with 4×8 ml volumes of sterile saline. The first lavage was retained in a separate tube for lactate dehydrogenase (LDH) and protein estimations. Untreated rats were lavaged to provide bronchoalveolar lavage (BAL) which was used in the experiments outlined later. Tubes were spun at 100 g for 5 minutes at 4°C, the supernatant was removed, the cell pellet from the first lavage was combined with the cells from the same lavage and resuspended in 1 ml phosphate buffered saline (PBS). Total cells were counted, cytocentrifuge smears were prepared for differential cell counts, and stained with Diffquick (Raymond A Lamb, London). Three hundred cells per slide were counted and the results expressed as the total number of neutrophils in the lung lavage.
TREATMENT OF PARTICLES WITH BAL AND BSA
A stock solution of bovine serum albumin (BSA, 0.5 mg/ml in saline) was prepared. The UfCB particles were suspended at concentrations ranging from 0.3–5 mg/ml in this solution and in BAL from control rats, and were briefly sonicated and incubated in a shaking water bath at 37°C for 1 hour. Samples were centrifuged at 145 g for 10 minutes and the supernatants were removed for protein estimations.
MEASUREMENT OF Fe(III)
The amount of Fe(III) released from the particles and combining with desferal at pH 4.6 and pH 7.2 was measured spectrophotometrically by comparison with a standard curve of FeCl3. One hundred microlitres of sample or standard were added to triplicate groups of wells on a 96 well plate and 100 μl 2 mM desferal was added to each well. The plate was immediately read on a Dynatec plate reader at 430 nm. Standards consisted of a range of concentrations of FeCl3 from 15.625 to 1000 μM diluted in buffer at the appropriate pH.
PROTEIN ASSAY
Biorad stock reagent was diluted 1/5 with distilled water and filtered. Two hundred microlitres of diluted reagent were pipetted into wells on a 96 well plate, triplicate groups per test and per sample. Five microlitres of sample were then added to the appropriate well, mixed, and incubated at room temperature for 15 minutes before reading on a plate reader at 450 nm. Standards consisted of BSA solutions ranging from 1 to 0.0625 mg/ml.
LACTATE DEHYDROGENASE ASSAY
Fifty microlitres of 0.75 mM aqueous sodium pyruvate (Sigma) solution containing NADH (Sigma) at a concentration of 1 mg/ml were pipetted into each well of a 96 well plate and incubated at 37°C for 5 minutes. A series of standards were prepared to give a range of dilutions representing 0–2000 Units/LDH/ml. Fifty microlitres of pyruvate/NADH solution gave a concentration of 2000 LDH Units/ml. Ten microlitres of test sample or appropriate standard were added to each well in triplicate groups and thoroughly mixed. The plate was incubated for exactly 30 minutes at 37°C. Fifty microlitres of 2,4-dinitrophenylhydrazine (Sigma) solution dissolved in 1 M HCl were added to each well and incubated at room temperature for 20 minutes. To develop the final colour, 50 μl 4 M NaOH were added to each well, the plate was mixed and allowed to stand for 5 minutes. The plate was read at 540 nm on an automatic plate reader.
CALCIUM MEASUREMENTS
Mono Mac 6 (MM6) cells were grown in continuous culture in RPMI medium (Sigma) containing 1 mM sodium pyruvate, 1 mM oxaloacetate, 1 μg/ml holotransferrin, non-essential amino acids, L-glutamine, penicillin/streptomycin, and 20% foetal calf serum (all Sigma). Cells were adjusted to 4×106 cells/ml, and centrifuged at 145g for 2 minutes. The medium was removed, and the cells resuspended in 1 ml PBS and transferred to an Eppendorf tube. The tube was centrifuged at 145 g for 2 minutes at 4°C, PBS was removed and the cells were resuspended in serum free Roswell Park Memorial Institute (RPMI) medium containing 23 mM Hepes buffer. Cells were loaded with 1 μg/μl Fura 2 AM (Sigma) in dimethyl sulfoxide (DMSO, 2 μl/ml cell suspension), the tube was wrapped in foil and incubated in a shaking water bath for 20 minutes at 34°C. After incubation, the tube was centrifuged at 145g for 2 minutes at 4°C, the medium was removed and replaced with 1.5 ml fresh serum free RPMI medium.
The Fura 2 AM loaded cells were transferred to a quartz cuvette containing a magnetic stirrer, placed immediately into a fluorimeter with heated block and basal fluorescence measurements obtained over 500 seconds. The fluorimeter was set up to give excitation wavelengths of 340 nm and 380 nm, emission 510 nm, and excitation and emission slit widths set at 5 nm. During the experiments, the cuvette temperature was kept constant at 37°C. Continuous readings were recorded at 1 second intervals. After 500 seconds, 100 μl appropriate treatment in RPMI medium was added to the cuvette. The experiment was allowed to run for a further 1500 seconds. Treatments consisted of CB or UfCB at 1 mg/ml (untreated or treated with desferal) to give a final concentration of 66 μg particles/1.5 ml medium, particle leachates diluted 1:10 in medium (to give the equivalent of 1 mg particles), or medium alone.
After the particle/leachate treatment, 7.5 μl 20 μM thapsigargin (Sigma) in DMSO was added to the cuvette and the experiment continued for a further 500 seconds. Twenty microlitres of 5% Triton solution was added to the cuvette to lyse the cells to give the maximum fluorescence (Rmax) and the experiment was continued for 500 seconds. To give the minimum fluorescence value (Rmin), 15 μl 0.5 M ethylene glycol tetraacetic acid (EGTA) in 3 M Tris buffer was added to the cuvette. The experiment was ended after a further 500 seconds. The ratio of the fluorescence measurements at excitation wavelengths of 340 and 380 nm were converted to calcium concentration values by the method of Grynkiewicz et al.18
DATA ANALYSIS
Data from all experiments were analysed with analysis of variance (ANOVA) with the Tukey multiple comparison test.
Results
IN VIVO MODEL OF LUNG INFLAMMATION
Particle instillation
The effect of instillation of 500 μg UfCB or CB with or without desferal treatment on neutrophil (PMN) recruitment 24 hours after instillation into the lung is shown in figure 1. The data show that after 24 hours of exposure to 500 μg particles there was a significant influx of PMNs only for UfCB compared with CB. Treatment with CB produced the same level of inflammation as the control group. Treating the particles with desferal before instillation produced no difference in PMN recruitment compared with the untreated particles. The mean number of PMNs increased about eightfold after treatment with UfCB or UfCB/desferal compared with the control group.
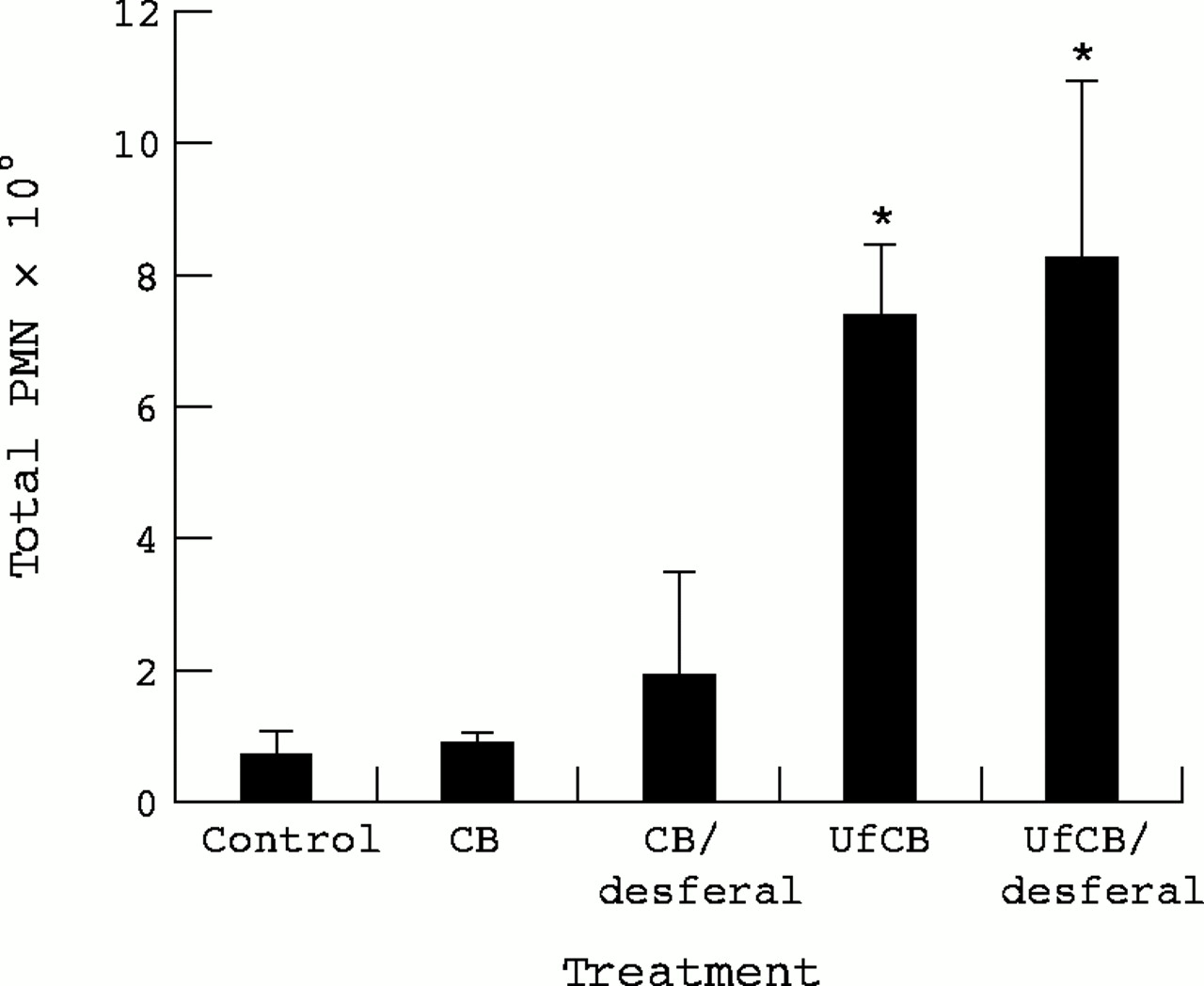
Neutrophil (PMN) recruitment in the lung 24 hours after instillation of 500 μg CB or UfCB with or without treatment of particles with desferal. Data are expressed as the mean (SEM) total PMN in BAL from three rats per treatment. (*p<0.05 CB v UfCB).
The LDH activity in BAL from particle treated rats is shown in figure2. There was no significant difference in LDH concentrations between control animals and CB treatment. The UfCB treatment produced an increase in LDH activity on average 769 Units/ml compared with 228 Units/ml for the controls, however, this increase was not significant (p>0.05). Desferal treatment of UfCB particles produced more LDH release than untreated particles and this effect was significantly different from the controls (p<0.05).
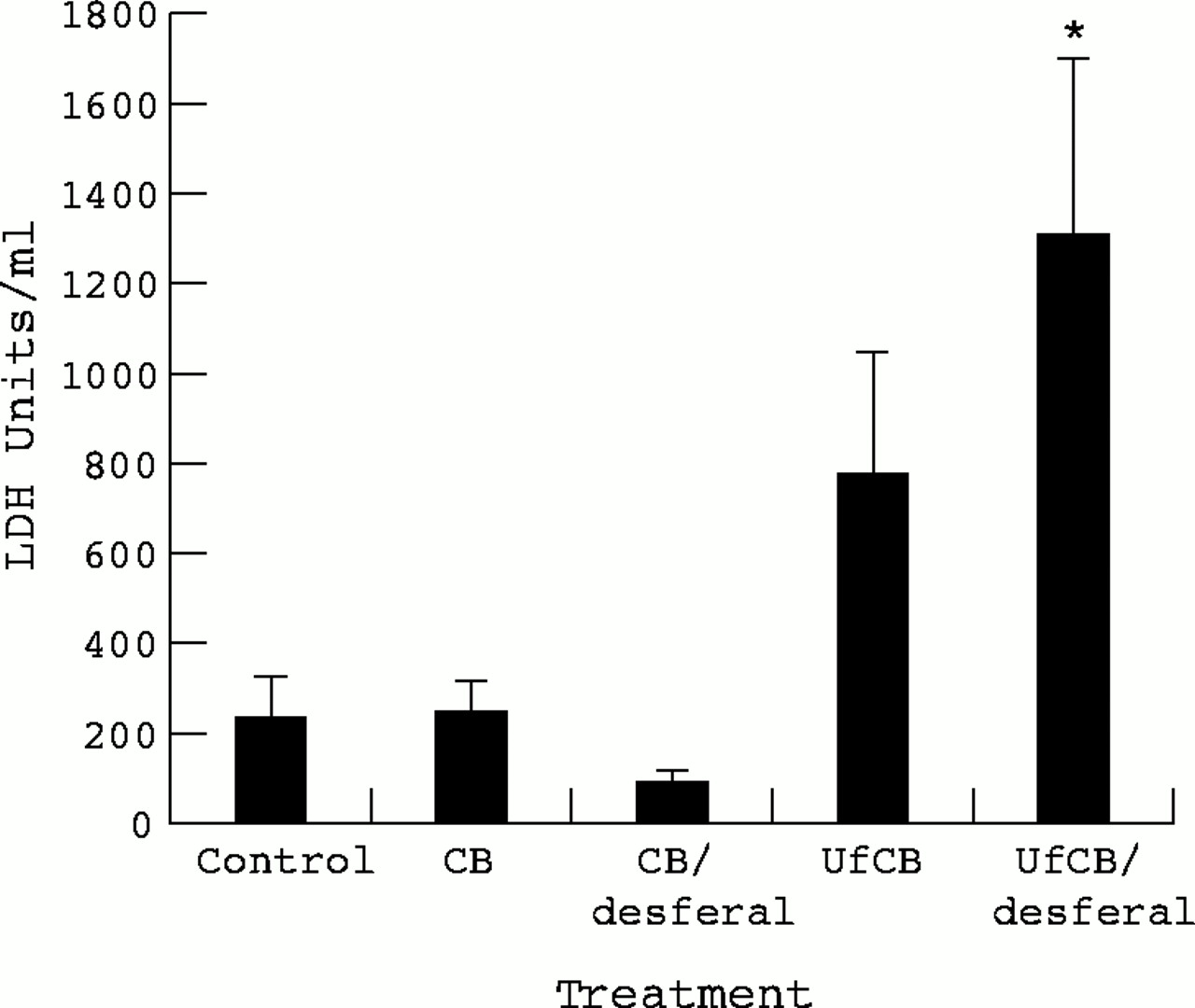
LDH activity in BAL 24 hours after instillation of 500 μg CB or UfCB with or without treatment of particles with desferal. Data are expressed as the mean (SEM) LDH units in BAL from three rats per treatment. (*p<0.05 Control v desferal treated UfCB).
Instillation of particle leachates
Leachates were prepared in both saline and mild citrate buffer. Citrate buffer was used as it is a mild chelator that would extract iron more readily from the particles. Instillation of either a saline or citrate leachate of CB or UfCB obtained from a mass of 1 mg particles produced no significant inflammation in the rat lung. Leachates from both particle types prepared in either saline or citrate buffer produced a similar number of PMNs in the BAL as the control, as shown in table 1.
Total neutrophils in BAL, 24 hours after instillation of 0.5 ml leachate, equivalent to a mass of 1 mg CB or UfCB prepared in saline or citrate buffer
IRON CONTENT OF LEACHATES
We attempted to measure the transition metal content of the leachates with desferal, but this was below the detection limits of the system, which was 15 nmol/ml. As we used 1 mg/ml we can conclude that this was less than 15 nmol of transition metal per mg particulate.
TRANSITION METAL ANALYSIS OF PARTICLE LEACHATES
Measurement of the transition metal content of particles with ICP-MS prepared in either saline or citrate buffer showed that there was no detectable iron in leachates of CB or UfCB in saline treated particles (table 2). Leaching into citrate, however, yielded detectable iron for both particle types, twice the amount was present in UfCB leachates than CB leachates (50 ng/mg particles for CB; 100 ng/mg particles for UfCB).
Transition metal content of CB and UfCB particles leached into saline or citrate buffer
DEPLETION OF PROTEIN FROM BAL AND BSA SOLUTION
Both particle types caused depletion of protein from BAL in a dose dependent manner (fig 3). This effect was most pronounced for UfCB, which depleted protein by 98% at the highest concentration of particles. The CB depleted protein in a similar manner, but by 45% for the highest concentration of particles (5 mg/ml). There was a significant (p<0.05) dose effect for UfCB particles and a significant difference between the particle types.

Protein content in BAL after CB or UfCB treatment. Data are expressed as a percentage of the control and represent the mean (SEM) of three separate experiments. There was a significant difference between particle type and a significant effect of dose for UfCB.
In keeping with the data produced with BAL, treatment of solutions of BSA as an indicator of protein with CB or UfCB resulted in depletion of protein in a dose dependent manner. These data are summarised in figure4. The reduction was more pronounced for UfCB. There was a significant (p<0.05) dose effect for UfCB particles and a significant difference between the two particle types.

Protein content in a solution of BSA after CB or UfCB treatment. Data are expressed as a percentage of the control and represent the mean (SEM) of three separate experiments. There was a significant difference between particle types and a significant dose effect of UfCB.
Ca2+ HOMEOSTASIS IN MM6 CELLS EXPOSED TO CB AND UfCB
The resting calcium concentration in MM6 cells was examined after addition of control particles and particles which had been treated with the iron chelator desferal (fig 5). For control cells, the mean (SEM) Ca2+ concentration was 68 (11.6) nM; after treatment with thapsigargin this increased to 390 (63.8) nM. There was some variation in the Ca2+ concentration of UfCB treated cells after thapsigargin stimulation between experiments—the lowest value was 391.5 nM and the highest was 1118.7 nM. Treatment with UfCB produced a significantly greater increase in cytosolic Ca2+concentration than CB (p<0.05) with particles both alone and after stimulation. Treatment of either CB or UfCB with desferal did not significantly alter the effects of either of these two particles on the resting Ca2+ concentration nor the response to thapsigargin compared with untreated particles.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Cytosolic Ca2+ concentration in MM6 cells after addition of CB and UfCB with and without treatment with desferal and after thapsigargin stimulation. Data are expressed as the concentration of Ca2+ in nM and represent the mean (SEM) of three separate experiments (p<0.05 CB v UfCB and CB plus desferal v UfCB plus desferal and after thapsigargin stimulation).
Leachates of both CB and UfCB prepared in acidic or neutral pH citrate buffer were used to investigate the effects of potential metal contaminants from the particles on calcium homeostasis in MM6 cells. The results of these experiments are summarised in table 3. There was an increase in the cytosolic Ca2+ concentration induced after treatment with thapsigargin for pH 4.6 leachate compared with pH 7.2 leachate although this effect was not significant. There was no significant difference in the resting cytosolic Ca2+concentration or the response to thapsigargin when CB and UfCB leachates were compared.
Cytosolic Ca2+concentration (nM) in MM6 after addition of leachate
Discussion
This study considers an important issue in the debate on the mechanism of lung injury caused by environmental particles (PM10). The components of PM10 include fine particles that tend to be secondary in formation comprising sulphate and nitrate, and ultrafine primary particles that are often aggregated, derived from combustion, and contain a carbon core. Studies have shown that transition metals can drive the proinflammatory effect of various types of environmental particles in vivo and in vitro.11We have shown that Edinburgh PM10 retrieved from TEOM filters can generate oxidants as shown by Fenton chemistry through the involvement of transition metals.13 We have also suggested that the ultrafine component of PM10 may mediate some of its toxicity.3 19 To fully understand the toxicity of PM10 it is necessary to dissect out the contribution that the components make to the total toxicity. Any ultrafine particles present could have adverse effects through transition metals associated with the surface, or ultrafine particles might have intrinsic toxicity regardless of transition metals. We used UfCB as a model ultrafine particle to find whether the increased inflammation they caused was, or was not, related to contaminating transition metals.
Toxicological studies of ultrafine TiO2 and CB5 8 20 show that ultrafine particles with no well documented transition metal content are highly inflammogenic. This has an important bearing as filter derived particle samples are likely to be highly biased toward the transition metal effect because the metals are soluble and in general filters are soaked in saline or something similar to release the particle sample. Indeed all the biological activity of some PM10 samples is in the soluble fraction.21 At the same time such filter samples may bias against the effects of ultrafine particles as these are likely to be highly adhesive and poorly retrieved from the filter matrix compared with soluble components. Our instillation data show that UfCB particles produced a significant neutrophil influx into the rat lungs compared with an equivalent mass of CB, as previously reported for a 125 μg dose.10 20
We used desferal as a chelator to study the role of transition metals and envisaged that there might be two effects of desferal on transition metals associated with particles. (1) Releasable or mobilisable transition metals would be drawn out of the particles and chelated. (2) Any transition metal that could not freely diffuse from the particle would be chelated on the particle. In either case, desferal treatment should show whether transition metals are involved in the proinflammatory effect of UfCB.
Earlier desferal treatment caused no significant effect on the inflammation caused by UfCB; CB had no significant inflammatory effect. As a measure of direct particle toxicity, LDH activity was measured in BAL after instillation. The only significant increase in LDH was caused by UfCB/desferal, but there was no significant difference between UfCB/desferal and UfCB with large variance in both. Therefore we place no biological significance on the difference between UfCB/desferal and UfCB and take it to be a statistical anomaly. The epithelium of the lung becomes more permeable during inflammation but we were unable to find any evidence of increased BAL protein as an indicator of epithelial permeability after UfCB or CB instillation (data not shown).
Reactive oxygen species have been strongly implicated in mediating the inflammation from ultrafine particles.22 The transition metal iron is important in redox cycling necessary to produce oxidants through Fenton chemistry and it has previously been shown, with a plasmid scission assay, that there is greater free radical activity associated with UfCB than with the same mass of CB.22However, we were unable to show any iron (III) in leachates of UfCB or CB, prepared in a neutral or acidic buffer when we used the desferal assay to measure iron. Iron was detectable in citrate leachates of both CB and UfCB with the ICP-MS method of iron detection. This is in keeping with the very low concentrations (ng/10 mg particles) of transition metal we reported previously in acid digests of CB and UfCB.10 When particle leachates prepared in citrate or saline were instilled into the rat lung, no inflammation was evident, suggesting that soluble factors or transition metals from the particles were not responsible for the lung inflammation found with particles alone.
Studies with carbon black particles at the cellular level have shown that UfCB but not CB decreases metabolic processes in epithelial cells through an oxidative stress mechanism.22 Stoneet al went on to describe the effects of CB and UfCB on calcium homeostasis in MM6 cells.14 The UfCB, but not CB, stimulated the entry of extracellular calcium and treatment with thapsigargin further increased the cytosolic calcium ion concentration compared with the controls or CB. In resting non-stimulated cells, calcium ions are sequestered into the endoplasmic reticulum, and plasma membrane Ca2+-ATPases maintain a low cytosolic calcium ion concentration which allows Ca2+ to act as an effective intracellular signalling molecule. Thus activation of transcription factors after exposure to ultrafine particles may be mediated in part by changes in calcium homeostasis and ultimately lead to the increased expression of proinflammatory genes. We have confirmed here the findings of Stone et al 14 that UfCB increased the resting cytosolic Ca2+ concentration and the size of the calcium response to thapsigargin was explained as an increase in the calcium current (ICRAC) activated by calcium release. In the present study, we set out to determine the role of transition metals in this phenomenon and we have extended the experiment to include CB and UfCB which had been treated with desferal and included leachates of CB and UfCB. Pretreated CB and UfCB had a similar effect to the untreated particles in the calcium experiments and leachates had no effect, suggesting that an effect mediated by iron or other soluble factors was not responsible for the enhanced calcium influx induced by UfCB.
In the absence of a clear role for transition metals in the inflammatory effects of UfCB we may look to the large particle number or surface area. Depletion of protein from BAL and solutions of BSA suggested that protein may be adsorbed on to the UfCB particle surface to a considerable extent. This may have an effect on epithelial cells by changing osmotic potential or changing the local conditions that may stress cells. The evidence of increased protein-surface interaction also raises the question of a role for complement. The alternative pathway of complement is activated when C3 binds to surfaces and this has been shown for asbestos and other particles.23 24 The ability to bind protein has been related to the ability to activate complement.23 The role of complement in the effect of ultrafine particles warrants further research.
It has been suggested by Oberdoster et al,5 that animals exposed to low concentrations of lipopolysaccharide (LPS) may become primed and on subsequent exposure to ultrafine particles may elicit an enhanced inflammatory response. This observation may help to explain why some people with underlying lung inflammation, such as asthma and chronic obstructive pulmonary disease, are susceptible to the adverse effects of particles. The calcium data presented here and reported previously14suggest that ultrafine particles may prime lung cells at the Ca2+ level through a mechanism not mediated by metal. This may result in enhanced inflammatory effects on subsequent exposure to proinflammatory cytokines stimulated by microbes or some other concomitant exposure to a pollutant. The converse may also be true—that pre-exposure to other pollutants or infection may prime lung cells to be hyperreactive to ultrafine particles, in terms of release of a proinflammatory mediator. The surface area or particle number of the UfCB provides the most likely explanation for the difference in inflammogenicity between UfCB and CB, and the UfCB could adsorb far more protein from lung lining fluid than the CB.
In conclusion our data show that, for one type of model ultrafine particle, the increased inflammation that it causes cannot be explained by transition metals nor other soluble factors released from UfCB. The study points out that ultrafine particles and transition metals may have independent proinflammatory effects in PM10. However, more studies on the association between transition metals and ultrafine particles in actual PM10 samples are required to fully elucidate the relative roles. Such studies are difficult to achieve but will be necessary to fully unravel the relative toxic potency of the components of the PM10 “cocktail”.
Acknowledgments
We acknowledge the assistance of the British Lung Foundation and the Medical Research Council. KD is the Transco British Lung Foundation Fellow in Air Pollution and Respiratory Health.